

































很多人都有这样的经历:买回来的家具看起来设计精巧,但装好之后却总有些地方不顺手——门关不严、抽屉卡顿。问题往往不在大件结构,而出在那些不起眼的连接部位。
PROTAC 分子的设计也有类似之处。一个典型的 PROTAC 由识别目标蛋白的 POI 配体、招募 E3 泛素连接酶的 E3 配体,以及连接两者的 linker(连接臂)组成。linker 看似只是把两个配体连在一起,实际上却同时影响三元复合物几何、构象分布、细胞渗透、代谢稳定性和代谢物性质,是决定候选分子能否继续推进的关键变量之一。
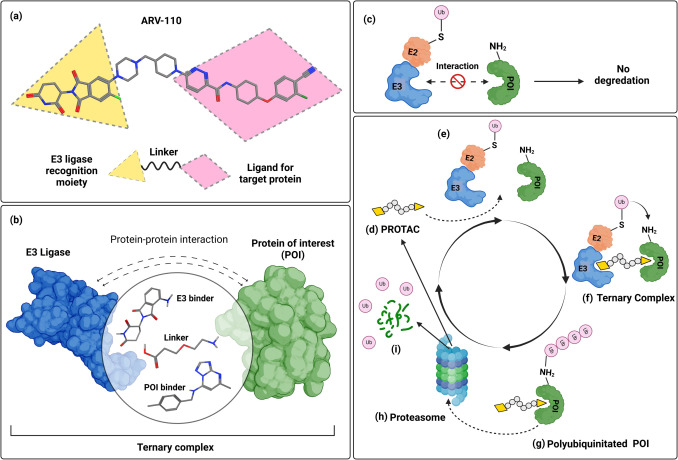
(a) PROTAC分子结构。 (b) 三元结构组成。(c) 如果没有 PROTAC,E2 和 E3 酶无法与 POI 相互作用,导致无法降解。(d) 引入 PROTAC 分子。(e) 形成三元复合物,使 E3 连接酶和 POI 能够接近。(f) POI 的泛素化。(g) 多聚泛素化标记 POI 以进行降解。(h) 转运至蛋白酶体进行降解。(i) POI 分解成肽,完成降解循环。PROTAC 被回收并准备形成新的三元复合物。
因此,linker 设计不是简单的“选一条长度合适的链”,而是一个需要同时处理药效、DMPK 与安全性约束的多参数优化问题。
一、Linker至少影响五个相互耦合的维度
1. 三元复合物的空间几何与动力学
PROTAC 发挥作用的关键,不只是分别结合目标蛋白和 E3 连接酶,而是形成能够驱动泛素化的“生产性三元复合物”。除总体稳定性外,复合物的协同性、形成与解离动力学、两个蛋白的相对取向,以及目标蛋白表面可被泛素化的赖氨酸是否处于合适位置,都可能影响最终降解效率。[1–3]
PROTAC 选择性诱导靶标降解的示意图
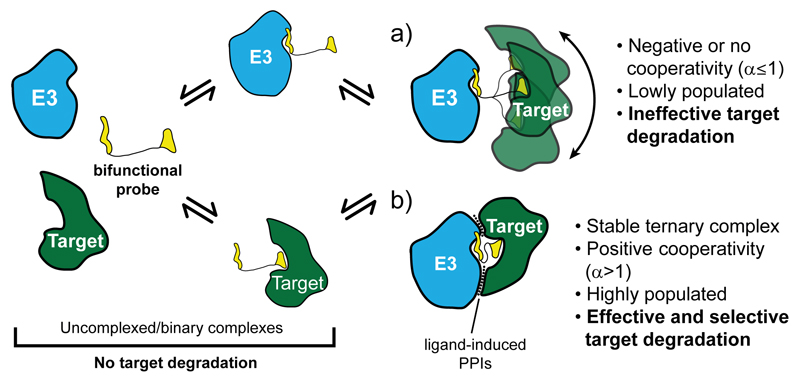
因此,linker 需要提供合适的距离、方向和构象约束。过短可能造成空间位阻,使两端配体无法同时结合;过长则可能增加构象熵,使有效取向在全部构象中的占比下降。但这并不是单调关系:不同 POI–E3 组合可能对应完全不同的最优长度和几何窗口。
2. 分子的构象自由度
柔性链段可以帮助分子适应不同蛋白表面,并探索更多可能的三元复合物姿态;但过高的构象自由度也会带来大量无效构象,并增加结合时的构象熵代价。
相对刚性的 linker 有机会减少无效构象、提高有利构象的占比,并使结构—活性关系更容易解释;但如果刚性片段锁定了错误方向,或限制了蛋白表面的适配能力,也可能显著削弱降解活性。
因此,Linker需要在“灵活”和“受控”之间找到平衡。
3. 细胞膜渗透
PROTAC 通常处于“超出五规则”(beyond rule of five,bRo5)的化学空间,常见特征包括分子量较大、极性较高、氢键供体/受体较多以及可旋转键较多,这些因素都会增加被动跨膜的难度。linker 的长度、极性、离子化状态和柔性会进一步改变这一平衡。
但二维结构参数并不能完整解释渗透性。一些 PROTAC 能够在不同介质中重新分配构象群,通过分子内氢键、π–π 相互作用或疏水折叠降低暴露的极性表面积,表现出“分子变色龙”特征。研究显示,linker 的化学性质与柔性对这类折叠构象的形成非常关键。[5,6]
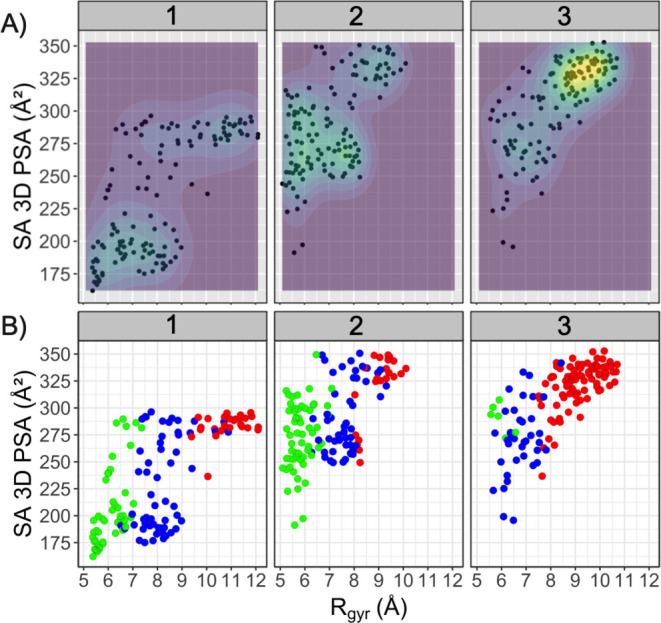
(A) 概述了由Rgyr和 SA 3D PSA定义的化学性质空间中,每个 PROTAC 的五个结构簇中 26 种构象子集的分布情况。性质空间中结构多样性更高的区域,即所选构象密度更高的区域,用黄绿色和等高线标出。(B) 化学性质空间中不同折叠构象的分布。折叠构象为绿色,半折叠构象为蓝色,线性构象为红色。请注意,某些构象占据相同的性质空间,因此在图 A 和图 B 中重叠显示。
4. 代谢稳定性
linker 往往是 PROTAC 的重要代谢区域之一,但并非所有分子都以 linker 为主要软点。具体代谢位置取决于完整分子的构象、电子环境、酶可及性和连接方式。常见反应可包括烷基或芳环羟基化、N/O-去烷基化、酰胺或其他连接键水解,以及不同形式的链段断裂。
较长的 PEG 样或线性烷基链通常会增加潜在氧化位点,但代谢稳定性并不只由链长决定。引入环状、不饱和或立体受限片段有时可以减少软点或改变酶识别方式;与此同时,也可能带来溶解度、渗透性或离子化方面的新问题。
5. 代谢物的药理与安全性风险
linker 发生断裂后,生成的片段不一定是完全失活的“废料”。POI 配体相关代谢物可能仍然结合靶蛋白,却无法招募 E3;E3 配体相关片段也可能保留一定结合能力。由于这些代谢物通常比母体分子更小,其游离比例、组织分布和清除速度可能明显不同。
已有研究表明,某些由 linker 不稳定性产生的 POI 结合型代谢物可以与完整 PROTAC 竞争靶点,从而削弱体内降解效果。[9] 因此,“母体药物减少”并不等于药理作用已经结束。
二、Linker 不是越长越好,也不是越短越好
在常规印象中,增加Linker长度似乎可以为两端配体提供更大的活动空间,让它们更容易分别找到目标蛋白和E3连接酶。然而,对PROTAC来说,过度延长Linker往往得不偿失。
当链段过长时,通常会出现几个问题:
第一,分子量继续上升,口服吸收和膜渗透更加困难。
第二,可旋转键增加,分子的构象数量迅速增多,真正有利于三元复合物形成的构象比例可能下降。
第三,代谢软点增多,Linker更容易发生氧化、去烷基化或断裂。
第四,两个蛋白之间的相对位置变得难以约束,虽然“够得着”,却未必能形成适合泛素化的几何关系。
另一方面,过短的 linker 可能造成配体或蛋白之间的空间冲突,也可能迫使三元复合物采用不利构象。
因此,更准确的设计目标不是寻找“最小有效长度”,而是寻找适合特定 POI、E3、连接位点和配体组合的最优几何窗口。这个窗口既包括端到端距离,也包括出射方向、局部刚性、可折叠性和构象群分布。
三、柔性有利于探索,刚性有利于约束,但二者都不是万能答案
在Linker优化中,另一个核心问题是柔性与刚性的平衡。
柔性 linker 常由 PEG、烷基链或混合链段构成,合成便利,也容易在早期快速探索 SAR。其代价是可旋转键较多、构象数量庞大,并可能引入多个氧化或去烷基化位点。
刚性化可以通过引入哌啶、环己烷、炔基、芳环或芳杂环等片段来降低部分构象自由度。潜在收益包括减少无效构象、改善代谢稳定性、改变折叠方式,并提高 SAR 的可解释性。但不同环系对极性、碱性、溶解度和渗透性的影响差异很大。例如,含可质子化氮的环状片段可能提高溶解度,也可能增加电荷负担;因此不能把“加环”简单等同于“更易成药”。
BTK PROTAC 的 6e→3e 优化案例可以说明这一点。6e 含有较长、柔性的 PEG linker,在小鼠肝微粒体中的半衰期约为 1.3 分钟;后续研究同时探索了刚性 linker 和 CRBN 配体,筛得 3e,其微粒体半衰期超过 145 分钟,BTK 降解活性也由 6e 的 DC50 约 41.9 nM 提高到 3e 的 7.0 ± 1.4 nM。[8]
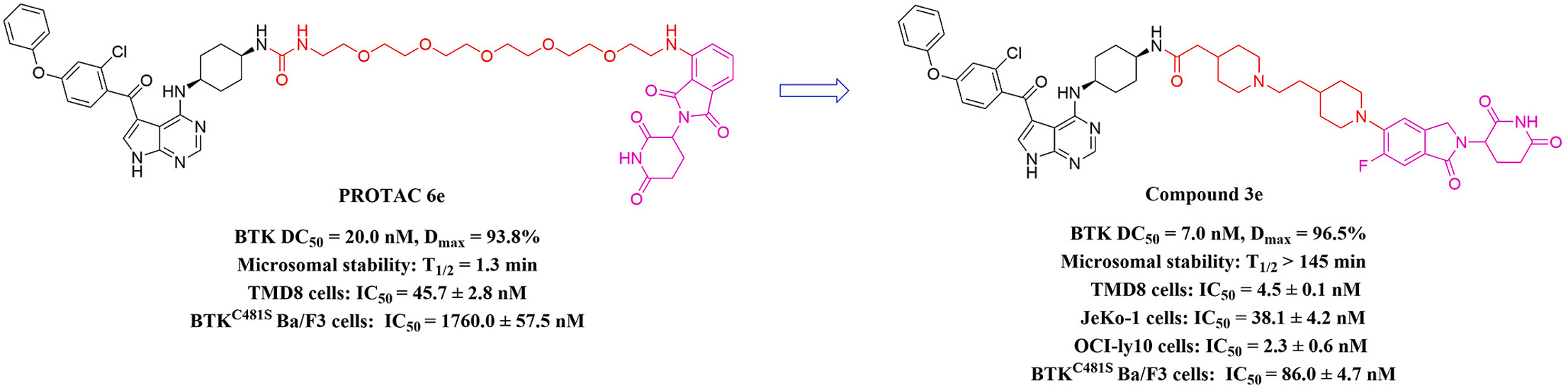
不过,这一案例不能被解读为“仅靠刚性化就能带来全部改善”,因为 linker 与 CRBN 配体均发生了优化。更准确的结论是:在合适的整体结构背景下,linker 刚性化可以成为同时改善药效与代谢性质的有效策略之一。
四、连接位点和出射向量,可能比链条本身更重要
谈到Linker优化,人们很容易把注意力集中在链长和化学组成上,却忽视了连接位点。
同一条 linker 连接在配体的不同位置,可能产生完全不同的降解活性、选择性和 DMPK 表现。连接位点会改变配体的出射方向、三元复合物的几何关系,也可能遮挡原本参与蛋白结合的关键相互作用。对 BET 降解剂的研究已经显示,改变 warhead 骨架或 linker 的连接向量,可以显著改变三元复合物识别和细胞降解活性。[4]
一个较合理的连接位点通常需要同时满足三点:
- 位于或接近溶剂暴露区域,尽量不破坏配体与蛋白的关键相互作用;
- 连接后仍能保持足够的 POI 与 E3 二元结合能力,并形成有利的三元复合物取向;
- 连接键及其邻近结构不存在明显的水解、氧化或去烷基化风险。
因此,在实际项目中,与其只围绕同一连接位点反复加长或缩短链条,不如并行建立多个 exit vector(出射向量)系列。很多时候,问题不是“链不够好”,而是“链从错误的方向伸了出去”。
五、Linker 会影响 PROTAC 如何穿过细胞膜
PROTAC 的作用对象通常位于细胞内,因此,足够的细胞暴露是实现降解的前提。这里存在一个常见矛盾:为了形成有利的三元复合物,分子可能需要一定长度和柔性;为了提高被动渗透,又希望减少暴露极性、可旋转键和电荷。
分子内氢键和环境依赖性折叠是解决这一矛盾的可能路径之一。需要强调的是,“变色龙效应”并不意味着分子一定在水相中完全展开、进入膜后才突然折叠。更准确的描述是:分子在不同介质中占据不同的构象群,低极性环境可能富集更紧凑、暴露极性更低的构象,从而有利于膜分配和跨膜。
因此,仅依靠分子量、cLogP、氢键数量和二维 TPSA 等静态参数,通常不足以预测 PROTAC 的渗透性。更有价值的评价应结合细胞渗透实验、溶液构象、分子内相互作用、离子化状态以及不同介质中的动态变化。
六、完整 PROTAC 不能被看成三个零件的简单相加
POI 配体、linker 和 E3 配体的单独数据可以提供设计线索,但不能直接外推出完整 PROTAC 的代谢和药代性质。三部分连接后,分子的构象、电子环境、空间遮挡、酶可及性和溶剂暴露都会发生变化。原本容易代谢的位点可能被隐藏,原本稳定的片段也可能因新的连接方式而出现软点。
一项针对 40 个 PROTAC 的人肝细胞代谢研究显示,完整分子的代谢行为不能由各组成配体的代谢结果简单预测,linker 的化学性质和长度会显著影响代谢易感性。[7]
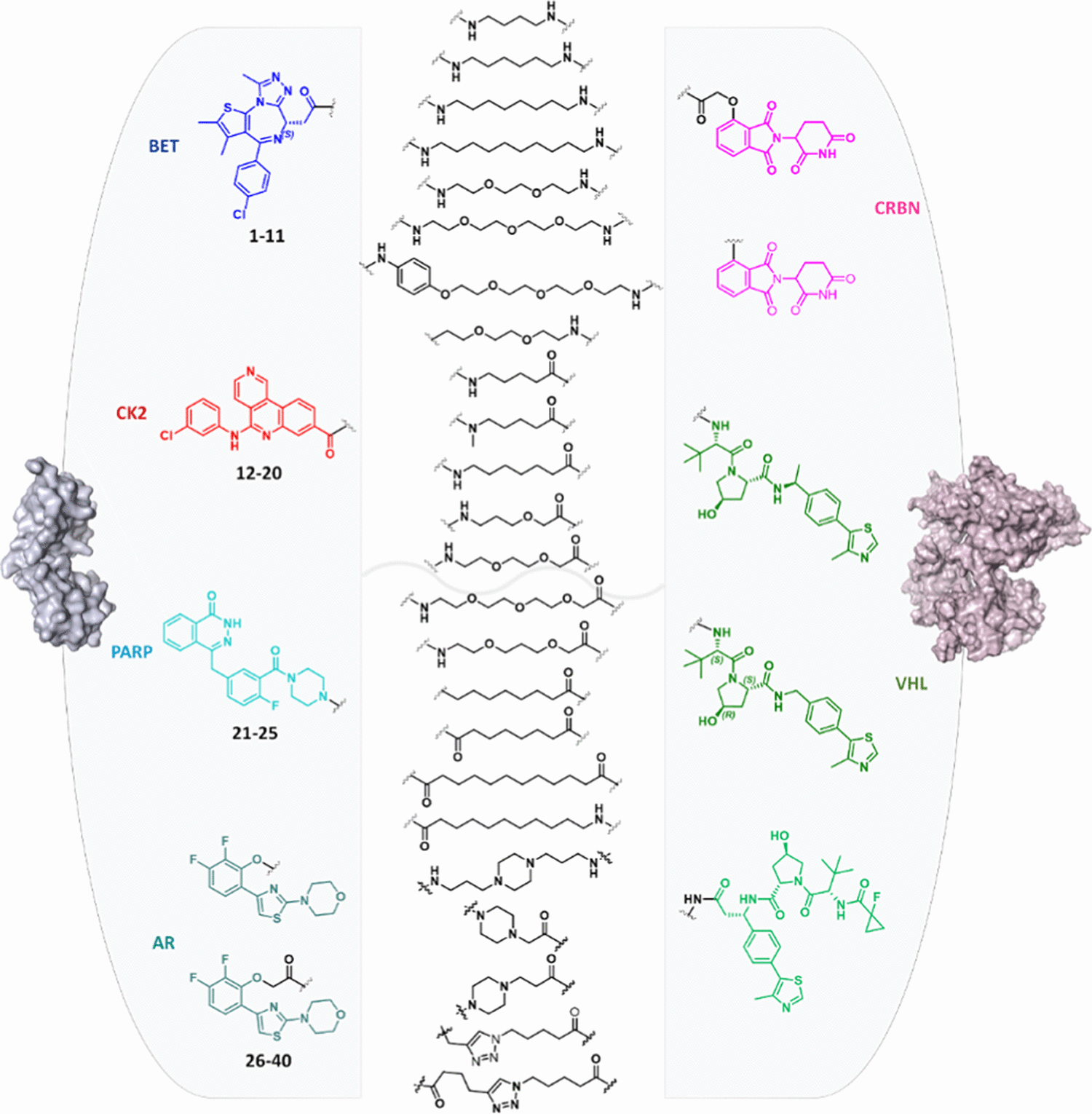
包含40个PROTAC的数据集
因此,PROTAC 应被视为新的、独立的化学实体,而不是几个已知模块的简单拼接。
七、Linker 断裂后,药效和安全性可能重新洗牌
linker 断裂并不是 PROTAC 独有的代谢方式,但对双功能降解剂尤其值得关注,因为断裂会把一个具有事件驱动作用机制的分子,转化为一个或多个可能仍具有占位型结合能力的片段。
从药效角度看,POI 配体相关代谢物可能继续占据靶蛋白结合位点,却不能招募 E3;如果暴露足够高,就可能表现为普通抑制剂,或与母体 PROTAC 竞争结合。2024 年报道的一项口服 ERα VHL-PROTAC 研究中,linker 代谢产生的靶点结合型代谢物被证实会竞争 ERα,并限制体内最大降解程度。[9]
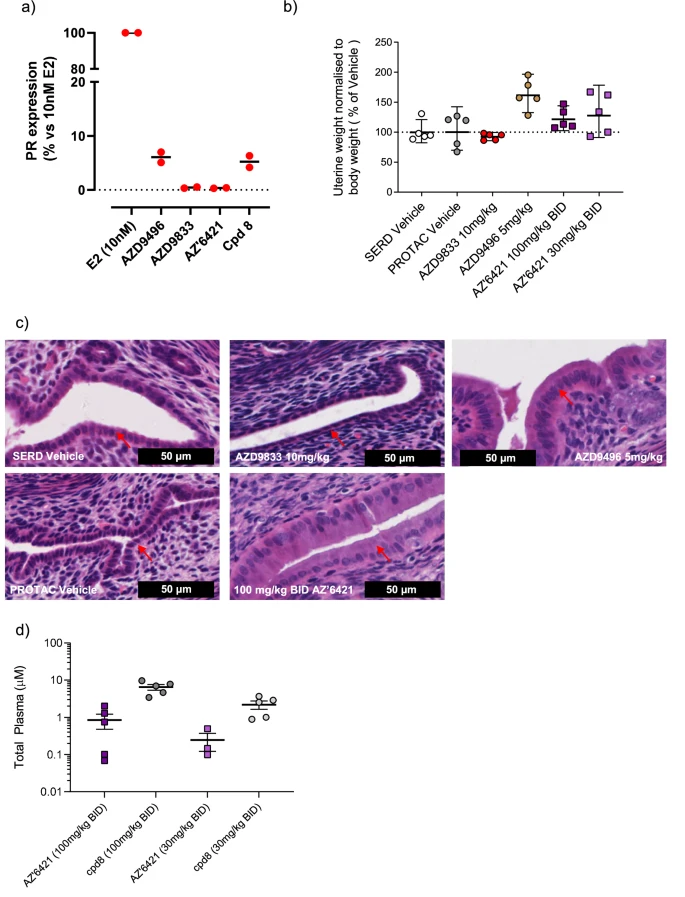
AZ'6421 和化合物 8 在体外和体内的 ER 激动作用的证据
从安全性角度看,主要代谢物的结合谱、游离比例、组织分布和物种差异也可能与母体不同。因此,对于存在明显链段断裂的候选分子,至少应回答以下问题:
1. 主要生成哪些代谢物,断裂发生在何处?
2. 这些代谢物在不同物种和不同生物基质中的形成比例是否一致?
3. 它们是否仍保留 POI、E3 或其他蛋白的结合能力?
4. 它们的暴露是否足以改变母体药物的 PK/PD 关系、疗效或安全窗口?
如果这些问题没有得到回答,仅监测母体药物浓度,可能无法完整解释体内药效不足、作用延迟或毒性信号。
八、Linker 优化应从“单指标修补”转向多参数平衡
linker 设计中最常见的误区,是发现一个问题后只修复一个指标。为提高溶解度而增加极性基团,可能降低细胞渗透;为提高渗透而增加脂溶性,可能加剧非特异性结合、低溶解度或代谢风险;为提高稳定性而大幅刚性化,也可能损害三元复合物的适配能力。
更现实的策略,是建立多参数评价框架,至少同步考察:
- 二元结合、三元复合物形成、协同性与降解动力学;
- DC50、Dmax、选择性以及钩状效应(hook effect);
- 水溶性、生理介质溶解度和聚集倾向;
- 细胞渗透、外排和细胞内游离暴露;
- 肝微粒体、肝细胞、血浆或全血稳定性;
- 主要代谢软点、代谢物鉴定和跨物种差异;
- 血浆蛋白结合、非特异性结合与组织分布;
- 母体及主要活性代谢物的 PK/PD 关系。
DMPK 评价不应等到降解活性优化完成后才开始。越早识别水解风险、氧化软点、低渗透、外排或高非特异性结合等问题,越有机会在同一轮 SAR 中同步解决,而不是在后期反复返工。
结语:Linker 设计是一门多目标平衡艺术
在 PROTAC 分子中,POI 配体决定“找谁”,E3 配体决定“请谁来”,而 linker 决定双方能否以合适的距离、方向和动力学方式相遇。
它影响三元复合物几何和协同性,调节分子的构象与折叠,参与决定细胞渗透和体内暴露,同时也可能成为代谢软点及活性代谢物的来源。
因此,优秀的 linker 并不追求某一个指标的极致,而是在降解活性、选择性、渗透性、溶解度、代谢稳定性和安全性之间找到适合具体项目的平衡。对 PROTAC 而言,这种平衡往往比“链长多少个原子”更接近真正的成药问题。
参考文献:
1. Gadd MS, et al. Structural basis of PROTAC cooperative recognition for selective protein degradation. Nature Chemical Biology. 2017;13:514–521. doi:10.1038/nchembio.2329.
2. Roy MJ, et al. SPR-Measured Dissociation Kinetics of PROTAC Ternary Complexes Influence Target Degradation Rate. ACS Chemical Biology. 2019;14:361–368. doi:10.1021/acschembio.9b00092.
3. Wurz RP, et al. Affinity and cooperativity modulate ternary complex formation to drive targeted protein degradation. Nature Communications. 2023;14:4177. doi:10.1038/s41467-023-39904-5.
4. Chan KH, et al. Impact of Target Warhead and Linkage Vector on Inducing Protein Degradation. Journal of Medicinal Chemistry. 2018;61:504–513. doi:10.1021/acs.jmedchem.6b01912.
5. Atilaw Y, et al. Solution Conformations Shed Light on PROTAC Cell Permeability. ACS Medicinal Chemistry Letters. 2021;12:107–114. doi:10.1021/acsmedchemlett.0c00556.
6. Poongavanam V, et al. Linker-Dependent Folding Rationalizes PROTAC Cell Permeability. Journal of Medicinal Chemistry. 2022;65:13029–13040. doi:10.1021/acs.jmedchem.2c00877.
7. Goracci L, et al. Understanding the Metabolism of Proteolysis Targeting Chimeras (PROTACs): The Next Step toward Pharmaceutical Applications. Journal of Medicinal Chemistry. 2020;63:11615–11638. doi:10.1021/acs.jmedchem.0c00793.
8. Chen S, et al. Discovery of novel BTK PROTACs with improved metabolic stability via linker rigidification strategy. European Journal of Medicinal Chemistry. 2023;255:115403. doi:10.1016/j.ejmech.2023.115403.
9. Hayhow TG, et al. Metabolism-driven in vitro/in vivo disconnect of an oral ERα VHL-PROTAC. Communications Biology. 2024;7:563. doi:10.1038/s42003-024-06238-x.
查数据,找摩熵!想要解锁更多药物研发信息吗?查询摩熵医药(原药融云)数据库(vip.pharnexcloud.com/?zmt-mhwz)掌握药物基本信息、市场竞争格局、销售情况与各维度分析、药企研发进展、临床试验情况、申报审批情况、各国上市情况、最新市场动态、市场规模与前景等,以及帮助企业抉择可否投入时提供数据参考!注册立享15天免费试用!

















 川公网安备51019002008863号
川公网安备51019002008863号 本网站未发布麻醉药品、精神药品、医疗用毒性药品、放射性药品、戒毒药品和医疗机构制剂的产品信息
本网站未发布麻醉药品、精神药品、医疗用毒性药品、放射性药品、戒毒药品和医疗机构制剂的产品信息
收藏
登录后参与评论
暂无评论