


































随着个体化用药理念的普及,越来越多的临床医生开始意识到:同一种他汀类药物、同样的剂量,为何有人血脂控制良好、有人却以横纹肌溶解综合征收场?为何二甲双胍在不同患者体内的降糖效果相差悬殊?答案,藏在细胞膜上一类被低估已久的蛋白家族里——溶质型转运体(Solute Carrier,SLC)。
与上篇文章介绍的ABC转运体不同,SLC转运体不直接消耗ATP,而是借助电化学电位差或离子梯度来驱动物质跨膜,属于易化转运体或次级主动转运体。目前已鉴定出逾300种SLC转运体,其中OATPs(有机阴离子转运肽)、OATs(有机阴离子转运体)、OCTs(有机阳离子转运体)、MATEs(多药/毒物外排泵)和PEPTs(类肽转运体)是与药物代谢关系最为密切的五大家族。

SLC转运体最重要的功能特征是:多数以摄取为主,介导底物从细胞外进入胞内;而MATEs则反其道而行,以外排为主。正是这种方向性的精巧配合,构成了肝、肾、肠、脑等各器官药物处置过程中一套完整的"进站—转运—出站"体系。
一、有机阴离子转运肽(OATPs):肝脏的"选择性入口"
OATPs的家族成员与分布
OATPs由SLC21/SLCO基因家族编码,人体中已确认11个成员,分属6个亚家族。其中,OATP1B1和OATP1B3高度特异性地分布于肝细胞窦膜,是绝大多数口服药物经肝首过效应的核心执行者;OATP1A2则兼顾小肠上皮顶膜、血脑屏障和肾远曲小管,参与药物在多个屏障部位的转运;OATP2B1分布于肝、肠、胎盘和心肌,调节小肠吸收与肝摄取的平衡。
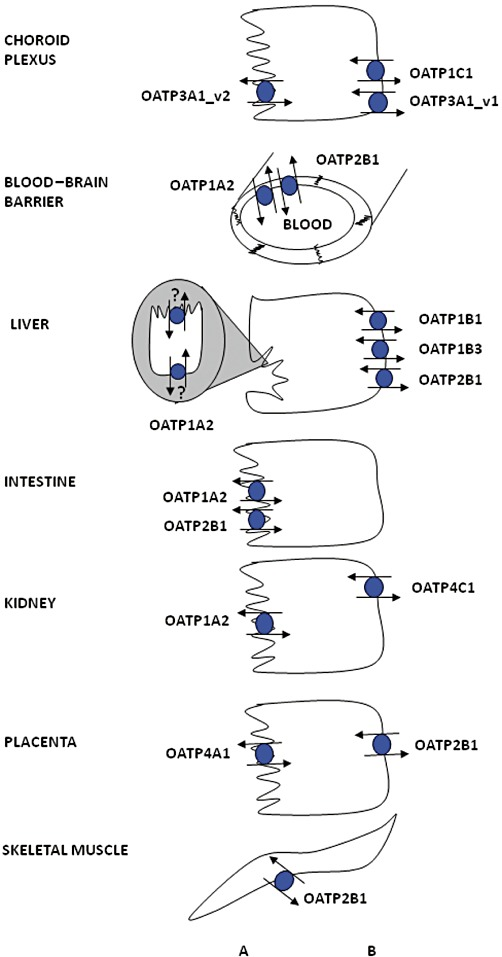
OATP在特定人类上皮细胞中的表达。已证实胆管细胞中存在OATP1A2的表达,但尚未确定其在特定细胞膜上的定位。(A) 顶端膜;(B) 基底外侧膜。(doi:10.1111/j.1476-5381.2011.01724.x)
OATPs的转运机制颇具特色——采用"跷板开关(rocker-switch)"构象转换模式,通过与细胞内HCO₃⁻或谷胱甘肽结合物进行物质交换,实现底物双向跨膜转运。这一过程是电中性的,不依赖Na⁺、K⁺或Cl⁻梯度,也不依赖膜电位和ATP,但对pH敏感——例如,酸性环境下OATP2B1的转运活性会明显增强,这也是葡萄柚汁能够干扰多种药物肠吸收的分子基础之一。
OATPs的主要底物与抑制剂
OATP1B1的内源性底物涵盖胆红素、胆汁酸、甾类激素结合物(如E₂17βG、雌酮硫酸酯、脱氢表雄酮硫酸酯)及前列腺素类;外源性药物底物则包括他汀类(辛伐他汀、普伐他汀、匹伐他汀等)、血管紧张素转化酶抑制剂、血管紧张素II受体拮抗剂等大量临床常用药物。
OATP1B3与OATP1B1的氨基酸相似度约80%,底物谱高度重叠,但胆囊收缩素是其特征性底物——这个细节常被用于区分二者的功能贡献。地高辛、多西他赛和紫杉醇也是OATP1B3底物,这一发现对肿瘤化疗中的药物相互作用评估具有重要价值。
OATP2B1的底物谱中,阿力吉仑(肾素抑制剂)是一个典型案例。合用OATPs抑制剂环孢素A后,因肝摄取被抑制,阿力吉仑的AUC和Cmax分别增加4~5倍和2.5倍;而葡萄柚汁通过抑制OATP2B1介导的肠吸收,可使阿力吉仑的AUC和Cmax分别降低61%和81%。同一底物,一个抑制剂让它暴露量飙升,另一个让它骤降——这一矛盾恰恰说明,转运体在摄取位点(肝)与吸收位点(肠)的方向性作用截然相反。
图源:美国食品药品监督管理局
OATP基因多态性:为什么同样的药剂量,效果却大相径庭?
目前已鉴定多种SLCO突变基因(SNPs),这些多态性可显著改变OATPs的表达和功能,进而影响药物的药代动力学及药效学。
以SLCO1B1为例,最常见的两种SNPs——c.521T>C(p.Val174Ala)和c.388A>G(p.Asn130Asp)——构成了4个功能不同的单倍型,即SLCO1B1*1A(野生型)、*1B、*5和*15。
他汀类药物与肌病风险是其中最典型的临床案例。以辛伐他汀为例:携带SLCO1B1c.521CC基因型者,其辛伐他汀酸的C_max和AUC约是521TT携带者的3.0倍和3.2倍;在521CC、521TC和521CC+TC携带者中诱发肌病风险的OR值分别为2.31、1.34和1.82,C等位基因携带者的肌病风险也显著高于T等位基因携带者。鉴于此,FDA已颁布了基于OATP1B1基因型的他汀类药物剂量建议。
OATPs介导的药物相互作用:三个必须记住的案例
案例一——吉非贝齐与西立伐他汀:吉非贝齐及其代谢产物吉非贝齐-葡萄糖醛酸苷是OATP1B1和CYP2C8的双重抑制剂。西立伐他汀既是OATP1B1的底物,又是CYP2C8的底物。两药合用后,西立伐他汀、其内酯代谢产物和代谢产物M1的AUC分别增加559%、440%和435%,横纹肌溶解综合征发生率大幅上升——这也是该药物最终退市的重要原因之一。
案例二——环孢素A与他汀类:环孢素A是OATP1B1、OATP2B1和OATP1B3的强效抑制剂,同时也抑制P-gp、BCRP和CYP3A4。合用环孢素A可使辛伐他汀AUC升高6~8倍、洛伐他汀升高5~20倍、阿托伐他汀升高6~15倍——这一数据足以说明,器官移植患者在接受免疫抑制治疗的同时,他汀类的剂量选择需要极为审慎。
案例三——利福平的"双面性":利福平既是CYP3A4的诱导剂,又是OATP1B1的抑制剂,其对阿托伐他汀的影响呈现出鲜明的时间依赖性——单次静脉注射利福平可使阿托伐他汀AUC增加7倍(OATP1B1抑制效应占主导),而口服利福平连续5天后AUC反而下降80%(CYP3A4诱导效应反超)。更有趣的是,这一相互作用的程度还取决于OATP1B1的基因型:口服600 mg利福平使阿托伐他汀在SLCO1B1 c.521TT、521TC和521CC携带者的AUC分别增加833%、468%和330%——基因型不同,"受伤"程度相差近3倍。
利福平化学结构
二、有机阴离子转运体(OATs):肾脏清除的三级主动接力
与OATPs主导肝脏摄取不同,OATs(由SLC22A基因家族编码)的主战场在肾脏。目前已鉴定OAT1~OAT10共10个成员,几乎遍布所有上皮细胞和内皮细胞。
OATs的转运机制颇为精妙,依赖二羧酸逆向交换驱动:OAT1和OAT3以α-酮戊二酸为交换底物,OAT2以琥珀酸或延胡索酸为交换底物,OAT7则交换短链脂肪酸(如丁酸)。维持细胞内高浓度二羧酸,最终需要Na⁺-二羧酸协同转运体(NaDC)和Na⁺/K⁺-ATP酶的接力支撑。正因如此,OATs介导的转运被定性为"三级主动过程"——这在转运体机制研究中是相当罕见的层级复杂性。
OAT1和OAT3主要表达于肾近曲小管细胞的基底膜,介导底物从血液侧摄取入肾小管细胞,随后经顶膜上的MATEs外排入尿液,完成经典的肾小管主动分泌通路。OAT3同时表达于脑毛细血管内皮细胞脑侧膜和蛛网膜上,参与底物从脑内和脑脊液的清除;OAT4则特异性表达于肾近曲小管刷状缘膜,主要介导底物从尿液的重吸收。
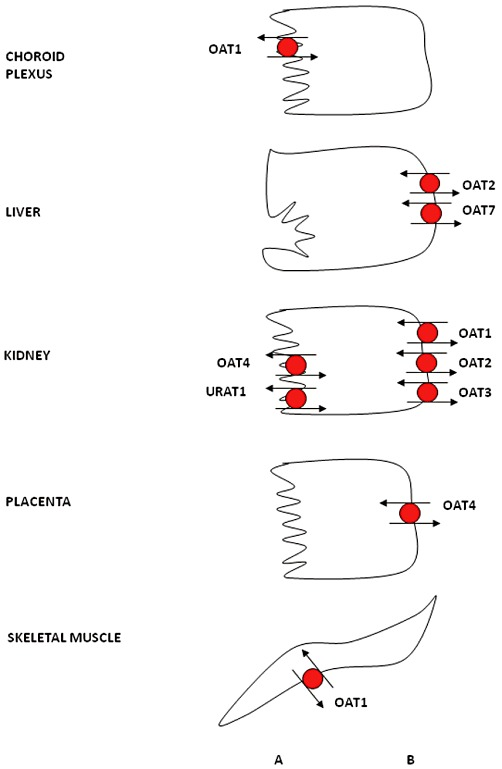
OATs在不同人类上皮细胞中的表达。根据啮齿动物数据推断,OAT1定位于脉络丛,OAT2定位于肝脏。(A)顶端;(B)基底外侧。
临床意义:从肾毒性到药物相互作用
OATs介导的药物相互作用有时是临床刻意利用的——最经典的案例是青霉素+丙磺舒。青霉素的肾排泄主要由OAT3介导,合用丙磺舒通过竞争性抑制OAT3,可延长青霉素的血浆半衰期、提高组织暴露浓度,延长疗效。这一策略在抗生素短缺时期曾是常规临床手段,如今依然有其实践价值。
然而,OATs的"摄取"功能也是一把双刃剑。头孢噻啶、阿昔洛韦、西多福韦等药物的肾毒性,正是因为它们被OATs大量摄入肾小管细胞后,在细胞内蓄积、造成毒性损伤。对策同样明确:合用丙磺舒等OAT抑制剂,通过竞争性抑制OATs介导的肾摄取,可防止药物在肾小管上皮细胞内蓄积,从而缓解肾毒性。丙磺舒与西多福韦的联合使用,已被推荐为标准临床治疗方案。
肾脏相关转运体还有一项容易被忽视的功能:介导肌酐的肾小管主动分泌(约占总清除的10%~20%)。当某些药物(如西咪替丁、甲氧苄啶、多替拉韦等)竞争性抑制OCTs、MATEs或部分OATs介导的肌酐清除时,血清肌酐水平会升高,而此时患者的肾小球滤过率(GFR)并未实际下降——这种‘假性’的肾损伤指标升高,属于单纯的药代动力学相互作用,临床解读时需注意甄别,以免误导诊疗决策。
脑OATs同样不容忽视。OAT3表达于血脑屏障和蛛网膜,介导底物从脑脊液和脑组织向血液侧清除。慢性肾病患者体内蓄积的尿毒素(如硫酸吲哚酚、马尿酸、吲哚乙酸等)竞争性抑制OAT3,导致神经递质代谢产物在脑内蓄积,这可能正是尿毒症脑病(记忆紊乱、精神错乱)的重要发病机制之一。
三、有机阳离子转运体(OCTs):从降糖药到神经精神领域
OCTs、OCTNs和CT2/OCT6同属SLC22A家族,均含有12个跨膜结构域(TMD)。OCTs是非Na⁺依赖性生电型转运体,驱动力来自跨膜电位差,介导有机阳离子药物的双向跨膜转运,不依赖pH——但底物的离子化程度会影响其与转运体的亲和力,pH降低往往增强跨膜转运效率。
OCT1主要在肝细胞窦膜表达,OCT2高度特异性分布于肾近曲小管基底膜,OCT3则作为神经元外单胺转运体广泛分布,尤以肌肉、肝脏、胎盘和心脏为主。
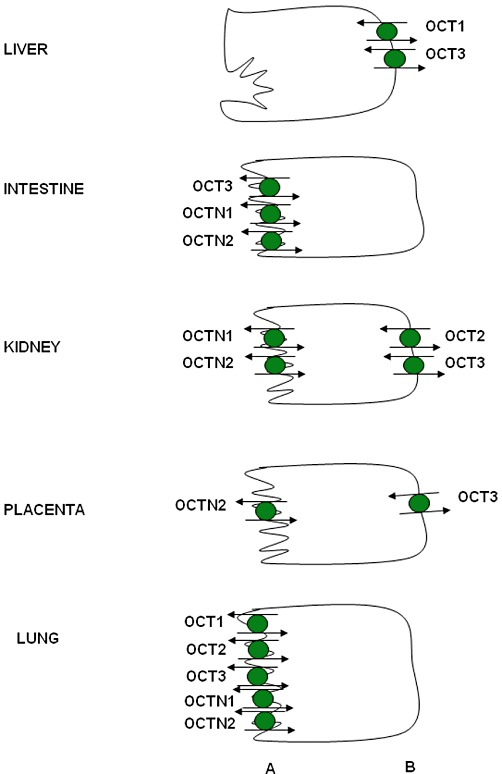
人上皮细胞中OCTs的表达。基于啮齿动物数据,得出OCTN1在肾脏中的定位结论。(A) 顶端;(B) 基底外侧。
二甲双胍与OCTs:一段关于疗效分层的故事
二甲双胍是OCTs在临床上最具代表性的底物,其降糖疗效的个体差异与OCT1/OCT2基因多态性密切相关。OCT1是二甲双胍的肝摄取转运体,已鉴定的功能丧失突变(Arg61Cys、Cys88Arg、Gly401Ser、Gly465Arg、Met420删除等)均可影响二甲双胍在肝内的分布与效应。携带两种OCT1功能丧失突变者,出现二甲双胍抵抗的风险约是未携带者的2倍以上。
肾脏方面,OCT2的rs316019(c.808G>T)多态性显著影响二甲双胍的肾清除率:GG>GT>TT。合用西咪替丁可降低二甲双胍的肾清除率,且这一效应在TT携带者中弱于GG携带者。OCT2基因突变还与二甲双胍导致高乳酸血症风险相关,TT携带者的风险高于GG携带者——这提示在特定基因背景下,二甲双胍的肾蓄积风险不可忽视。
此外,一些药物如西咪替丁、艾沙康唑、雷诺嗪、甲氧苄啶和凡德他尼等尽管可升高血清肌酐水平,但肾功能未见病理性改变,这归因于对肾OCT2活性的抑制——和前述OATs的情况类似,这是一种功能性而非器质性的肌酐清除率变化,临床解读时需注意甄别。
OCTs与脑功能:一个被忽视的神经药理窗口
OCT3广泛表达于脑的多个区域,作为神经元外单胺转运体,参与多巴胺、5-羟色胺和去甲肾上腺素的突触外清除,对脑内单胺环境的稳态维持发挥重要作用。OCT3基因突变往往与精神兴奋剂敏感性增加和焦虑行为相关,临床研究还发现OCT3基因突变与强迫症存在关联。OCT2也参与焦虑和抑郁等行为的调节,敲除Oct2的小鼠显示出焦虑样行为改变,且对抗抑郁药文拉法辛的应激应答发生变化。OCT2和OCT3已被提出可作为治疗心理障碍的潜在药理靶点。

四、多药/毒物外排泵(MATEs):肾脏分泌的"最后一道关卡"
MATEs(由SLC47A家族编码)通过与H⁺的反向交换实施有机阳离子的外排转运,其驱动力是顶膜内外的质子梯度([H⁺]in>[H⁺]out),而维持这一质子梯度依赖Na⁺/H⁺交换器或ATP-H⁺泵。人体中,MATE1表达于肾小管上皮细胞刷状缘膜和肝细胞胆管侧膜,MATE2-K则几乎仅见于肾小管刷状缘膜,是肾特有的转运体。MATE1和MATE2-K共同介导有机阳离子药物由肾小管上皮细胞分泌入尿液的过程,与基底膜侧的OCTs和OATs形成功能接力。
MATEs与顺铂肾毒性:转运体特性决定毒性命运
铂类药物的肾毒性差异,是MATEs临床意义最生动的诠释。顺铂和奥沙利铂都是OCT2的底物,均可经基底膜被摄入肾小管细胞;然而,奥沙利铂同时也是MATEs的高效底物,能通过顶膜迅速外排入尿液;而顺铂的MATEs底物亲和力远弱于奥沙利铂,导致其在肾小管细胞内大量蓄积,进而引发强肾毒性。卡铂和奈达铂既不是OCT2底物,也不是MATEs底物,因而绕过了这一蓄积机制,肾毒性显著低于顺铂。
动物实验进一步验证了这一机制:Mate1⁻/⁻小鼠中顺铂的肾毒性和肾蓄积显著高于野生型,合用乙胺嘧啶(强MATEs抑制剂)也可增加顺铂肾毒性。值得注意的是,抑制OCT2(或OCT2缺陷)反而降低顺铂肾蓄积、减轻肾毒性——这说明顺铂肾毒性的净效应,实际上是OCT2(摄入)与MATEs(外排)抑制作用的整合结果。
西咪替丁的"身份再认识"
西咪替丁长期以来被认为是OCT2抑制剂,但更精确的研究表明,其对MATE1和MATE2-K的抑制Ki值(分别为1.1~3.8 μmol/L和2.7~6.9 μmol/L)接近临床治疗剂量下的游离血药峰浓度(2.03~5.20 μmol/L),而对OCT1和OCT2的抑制Ki值则远高于临床血浆浓度(101~275 μmol/L和95~207 μmol/L)。这意味着,西咪替丁与二甲双胍等药物发生的肾相互作用,主导机制是MATEs抑制而非OCTs抑制——这一认知的更新,对药物相互作用的风险评估具有实质性意义。
五、类肽转运体(PEPTs):口服生物利用度的"主动推手"
PEPTs(PEPT1和PEPT2,由SLC15A家族编码)是质子驱动的转运体,利用顶膜质子电化学梯度介导肽类物质的跨膜转运。PEPT1主要表达于十二指肠至回肠的肠黏膜上皮细胞顶膜,容量高、亲和力低,是众多口服药物实现有效肠吸收的关键通道;PEPT2主要表达于肾近曲小管S3段刷状缘膜及蛛网膜上皮细胞顶膜,容量低、亲和力高,主导肽类药物的肾小管重吸收与脑脊液清除。
PEPT1的底物谱相当宽泛:β-内酰胺类抗生素(如头孢羟氨苄)、抗病毒药(如伐昔洛韦和更昔洛韦)、L-多巴前体、黏菌素类抗生素、5-氨基乙酰丙酸、以及部分血管紧张素转化酶抑制剂(如佐芬普利和福辛普利)均经此途径吸收。伐昔洛韦和pomaglumetad methionil是前药策略的典型范例——经肠PEPT1吸收后迅速水解为活性代谢产物阿昔洛韦和LY404039,兼顾了口服生物利用度与活性成分的靶向递送。
糖尿病与肠PEPT1:一个被忽视的吸收干扰因素
有文献报道,血糖控制不良的糖尿病患者体内氨苄西林血药浓度低于正常人,与氨苄西林的口服吸收降低相关。动物实验进一步证实:给糖尿病大鼠灌胃头孢氨苄和伐昔洛韦后,血浆中头孢氨苄和阿昔洛韦的浓度显著降低,这与大鼠肠Pept1蛋白表达受损密切相关。换言之,对于需要长期口服β-类抗生素或抗病毒药物的糖尿病患者,血糖控制状态本身可能构成影响药物吸收效率的隐性变量。
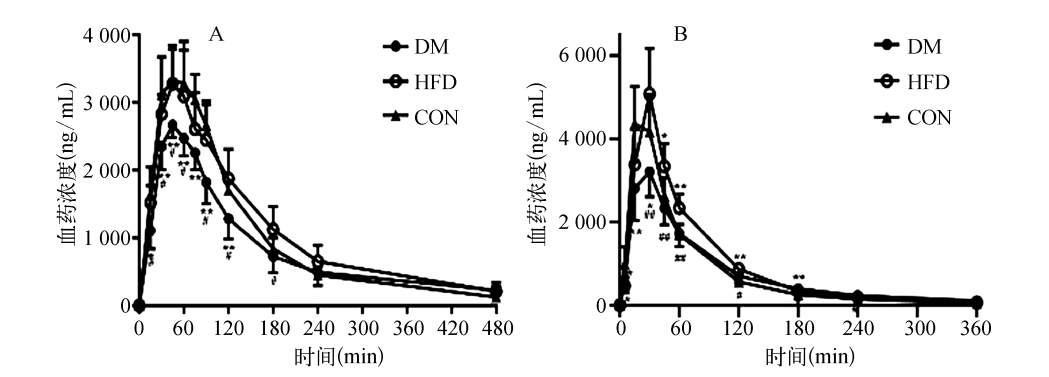
糖尿病(DM)大鼠、高脂饲养(HFD)大鼠和对照(CON)大鼠分别灌胃给予头孢氨苄(5mg/kg)或伐昔洛韦(18.4mg/kg)后的血浆中头孢氨苄(A)和阿昔洛韦(B)浓度-时间曲线
值得一提的是,PEPT1转运是质子依赖性的,pH对其活性影响显著——在pH 6.0时,Gly-Sar的转运活性比pH 7.5时高5倍。这意味着胃肠道局部pH异常(如质子泵抑制剂使用、胃酸分泌亢进或不足)均可间接影响经PEPT1转运的药物吸收效率,这一维度的药物相互作用在临床实践中值得关注。
脑PEPT2:神经保护的分子机制
PEPT2主要表达于蛛网膜上皮细胞顶膜,介导肽类物质从脑脊液向血液侧的定向转运,是维持脑内神经肽内稳态和清除神经毒性代谢产物的重要机制。5-氨基乙酰丙酸是卟啉类化合物前体,同时也是PEPTs底物,可诱发急性卟啉病。Slc15a2⁻/⁻小鼠脑脊液5-氨基乙酰丙酸浓度比野生型高5倍,皮下注射5-氨基乙酰丙酸后30 min和240 min,敲除小鼠脑脊液中的浓度分别比野生型高8倍和30倍——这直接证实了脑PEPT2在神经毒素清除中的保护作用。临床试验也显示,携带SLC15A2*2/2纯合子的儿童表现出更弱的运动灵巧性和记忆能力,提示PEPT2功能变异可能影响神经发育。
六、结语
回顾上述内容,SLC转运体家族对药物处置的影响可以归纳为三个核心维度:
物种差异造成的预测困境。OATPs的保守性差,小鼠Oatp1a4在肝脏和脑微血管内皮细胞上的表达与人OATP1B1、OATP1A2存在显著差异。在动物模型中获得的转运体相互作用数据,往往不能直接外推至人体——这是新药开发中一个经反复证实的陷阱,在药物转运体研究设计中须保持足够的审慎。
遗传多态性驱动的个体化用药需求。以SLCO1B1为代表,SLC转运体的功能性SNPs可使同种药物在不同基因背景患者中的血浆暴露量相差数倍,进而导致截然不同的疗效与毒性结局。FDA已将OATP1B1基因型纳入他汀类药物剂量建议,这是精准医学从理论走向临床的具体体现,未来有望延伸至更多药物类别。
转运体介导的药物相互作用是临床安全的隐性风险。无论是他汀类+吉非贝齐、他汀类+环孢素A,还是顺铂与MATEs抑制剂的叠加,背后都是转运体底物谱与抑制剂谱的交叉重叠。系统掌握各转运体的底物谱、抑制剂谱,以及临床血药浓度与抑制Ki值的定量比较,是合理预测和规避此类风险的科学基础。
SLC转运体的研究仍在快速推进。从基于PEPT1的前药设计、到靶向OATP1B1的肝特异性递药、再到通过调控MATEs减轻铂类肾毒性——转运体正在从"药代动力学的影响因素"升级为"药物设计的主动工具"。这场发生在细胞膜上的分子级博弈,终将以更精准的用药方案,造福每一位患者。
扩展阅读:
1. 2026年值得关注/追踪的抗体偶联药物:T-DXd、Dato-DXd、双抗ADC谁主沉浮?
2. 揭秘肿瘤多药耐药“元凶”:P-gp/BCRP/MRP三大ABC转运体如何构建癌细胞“防火墙”?
3. 突破溶解度瓶颈:早期药物开发中的粒径控制与纳米化策略解析
查数据,找摩熵!想要解锁更多药物研发信息吗?查询摩熵医药(原药融云)数据库(vip.pharnexcloud.com/?zmt-mhwz)掌握药物基本信息、市场竞争格局、销售情况与各维度分析、药企研发进展、临床试验情况、申报审批情况、各国上市情况、最新市场动态、市场规模与前景等,以及帮助企业抉择可否投入时提供数据参考!注册立享15天免费试用!


















 川公网安备51019002008863号
川公网安备51019002008863号 本网站未发布麻醉药品、精神药品、医疗用毒性药品、放射性药品、戒毒药品和医疗机构制剂的产品信息
本网站未发布麻醉药品、精神药品、医疗用毒性药品、放射性药品、戒毒药品和医疗机构制剂的产品信息
收藏
登录后参与评论
暂无评论